 EasySep™小鼠TIL(CD45)正选试剂盒
EasySep™小鼠TIL(CD45)正选试剂盒




肠类器官培养
肠类器官培养
- Document # MRDX20358
- Version 1.0.0
- 4/1/15
肠上皮细胞
哺乳动物肠道的腔表面由一层快速自我更新的上皮细胞组成,在5天内即可被新生细胞完全更替1。 上皮分为两个不同的区域,分别是隐窝区和绒毛区。隐窝区位于肠上皮最底部,与基底膜直接接触。隐窝中含有肠干细胞(ISCs)。 相反,绒毛区位于肠上皮的顶层表面,含有多种已分化的细胞类型1。
肠上皮主要有5种细胞类型。增殖的干细胞位于隐窝的末端区域,两侧是Paneth细胞,这种细胞分泌的细胞因子有助于维持干细胞微环境。Paneth细胞还负责分泌抗微生物多肽1。 干细胞对称分裂,根据分裂干细胞在隐窝内的具体位置,要么更新隐窝基底部的干细胞群,要么通过成为可快速分裂的隐窝短暂扩充细胞群的一部分开始分化。短暂扩充细胞向上朝顶部绒毛处移动,并最终在那里分化为具吸收功能的肠细胞、分泌激素的内分泌细胞或分泌粘液的杯状细胞。成熟上皮细胞沿绒毛向上移动,最终从绒毛尖端向管腔内片状脱落。
研究人员最近才通过谱系跟踪实验明确鉴定出:成熟的肠增殖干细胞群位于隐窝底部,由与Paneth细胞相互交织的柱状细胞构成2。 这些细胞表达LGR5(WNT信号通路中的一种蛋白),可以衍生为成熟肠道的所有细胞类型。 进一步的研究表明,隐窝基底部以外的干细胞,特别是隐窝+4位置中的细胞,在肠损伤时会还原成多能干细胞3。 这些发现证明在肠组织中干细胞和祖细胞的可塑性对于在肠腔恶劣环境内较严酷环境下维持稳态是十分必要的。
LGR5+干细胞保持自我更新和再生为肠上皮的能力,主要是因为它们处于干细胞微环境内4。 肠微环境已经被充分表征,研究显示其内含高WNT和上皮生长因子(EGF)组成的空间梯度,并可抑制骨形态发生蛋白(BMP)信号5。 现已针对这一确定的微环境研究构建了中性漂移模型。在该模型中,干细胞在空间受限的微环境中对称分裂。这种增殖必然导致一些干细胞从微环境中被去除,余下的干细胞则保持在隐窝基底上的位置,从而维持了它们的“干性”6。 随着时间推移,当相邻干细胞的子代细胞从微环境中逐渐被去除,某一既定隐窝内的干细胞将转向具有克隆性。
对有关肠上皮细胞谱系和功能,以及该组织可以不断更新的特性的掌握,使其成为一个非常具有吸引力的模型系统,可将其应用于研究上皮组织和成熟干细胞的一般特征,以及肠道相关疾病和药物开发的具体特点。
肠类器官历史
通过对小鼠模型和永生化细胞系(如:CACO-2腺癌细胞系)进行遗传操作的研究,我们已经能够充分掌握了肠的功能,包括组成上皮组织各种细胞类型的作用7。 得益于对这部分知识的了解,后来又开发出以尚未发生转化的原代细胞为起始样本所构建的肠上皮离体、长期模型。早期为构建这一培养系统,曾尝试创造了一些模型,在这些模型中,肠干细胞可以瞬时分裂,但会在数周内失去增殖能力8,9。
最早能够促进肠干细胞长期生存的培养系统是由Calvin Kuo博士的团队发表的10。 过培养含有上皮和间充质细胞的新生鼠的肠片段,Ootani等设计了一种气•液界面模型,在添加了胎牛血清后,生成隐窝状的结构,其中含有成年小鼠的所有主要肠细胞类型。如果饲育得当,这些培养物可以维持一年以上10。
同年,Hans Clevers博士的团队发表了另外一种用于培养肠隐窝的技术:以来源于成年小鼠肠组织并经过挑选的单个LGR5+细胞作为起始材料,建立体外组织培养模型。该系统取代了气•液界面,通过在半固体、富含层粘连蛋白/胶原蛋白的Matrigel®半球体中模拟细胞外基质来培养隐窝。结合使用半固体基质,该细胞培养基可以模拟隐窝的干细胞微环境,并长期培养这些细胞。相比于Ootani等10描述的囊状结构,该模型生成的“类器官”包含中心腔,及环绕于周围的代表肠隐窝的“芽”(图1)。这些隐窝状区域在功能上相当于已发育成熟的肠,因为分裂的LGR5+干细胞与Paneth细胞相互交错,且位于隐窝的基底部。因此,这些类器官具有高度生理相关性的表型。在Clevers团队开发的系统中,LGR5+干细胞分裂生成自我更新的干细胞群,以及终末分化为肠细胞、肠内分泌细胞或杯状细胞的细胞。终末分化细胞最终从绒毛状区域中排入管腔,即模仿了成熟肠上皮的生理更替周期。以这种方式培养的类器官可以通过周期性细胞解离和传代被无限期地维持培养(图2)5。 这一进步为相关研究建立了一种新型的模型系统,既具有体外研究的优势,即:易于培养,以及对在体内培养模型中起到关键作用的特定因子具有可控性,又可获得在体内模型中表现的多种生理相关特征,因此将两方面的优势进行了完美地结合。
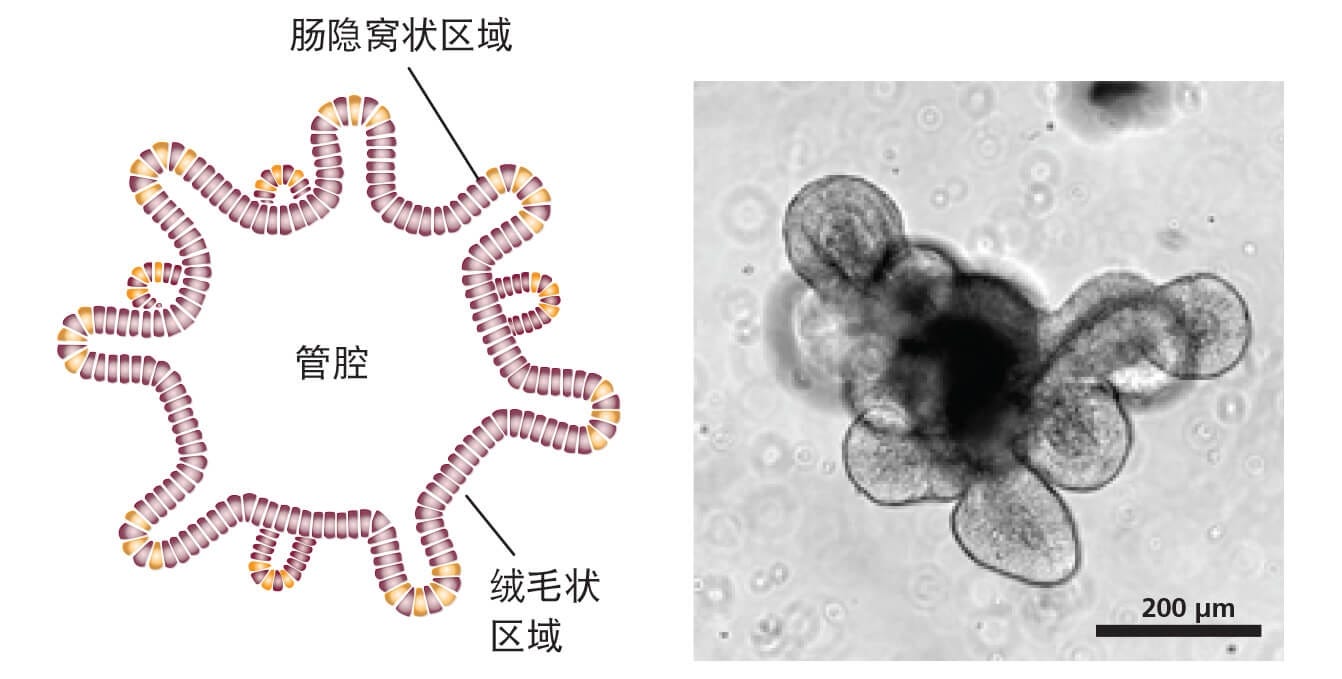
图1. 肠类器官形态
成熟的肠类器官的示意图(左)和成熟小鼠肠类器官(第5天)的明视图像(右)。中心腔由单层上皮和“萌芽”的隐窝状区域所围绕。
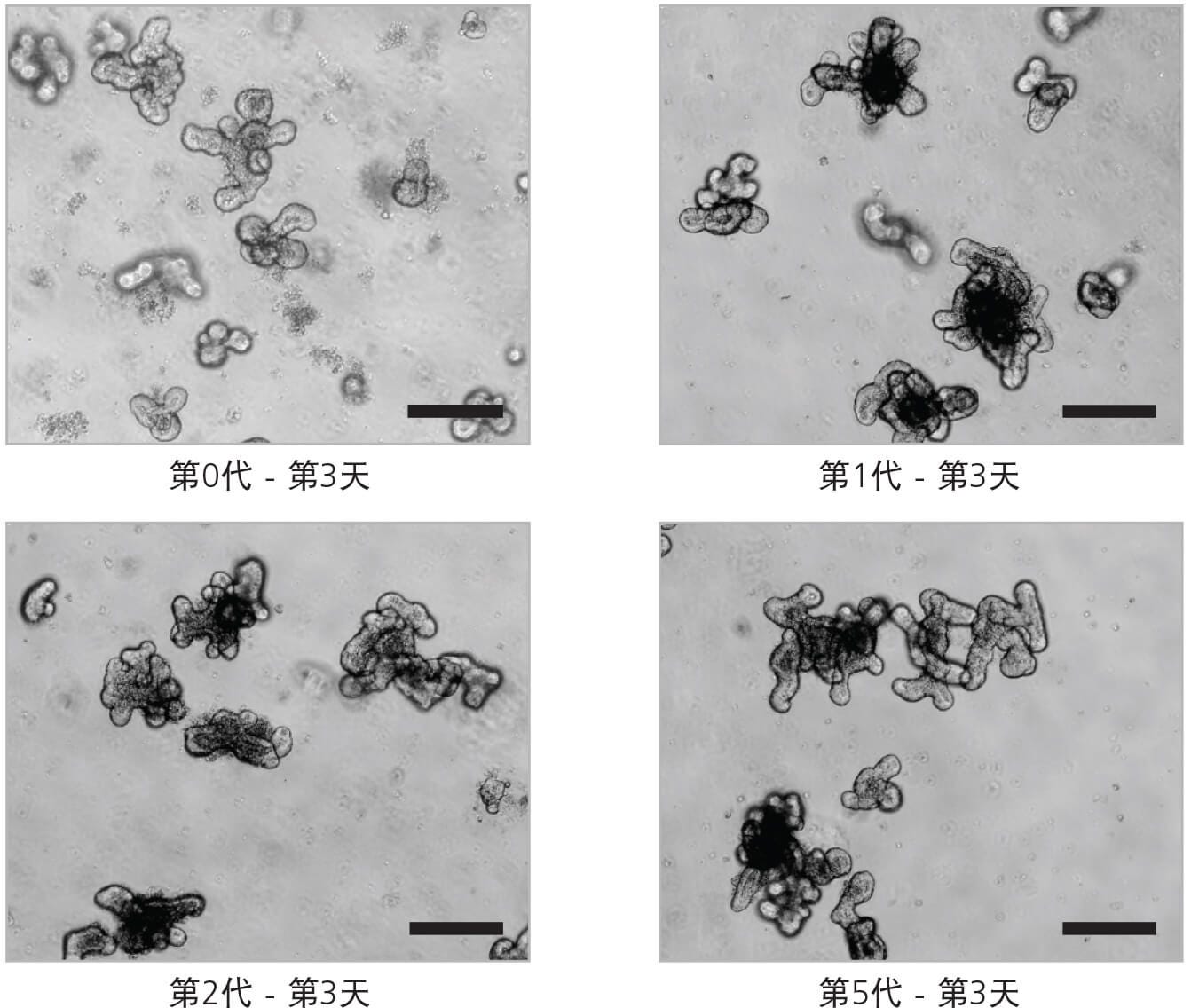
图2. 肠类器官的传代
源于小鼠初始隐窝的肠类器官典型示意图:第0代 - 第3天、第1代 - 第3天、第2代 - 第3天和第5代 - 第3天显示一直保持“萌芽”表型(比例尺=200 μm)。
培养肠类器官
开发更佳一致性和稳定性的培养基,以用于维持肠类器官隐窝状结构的干性,对于成功进行类器官培养是至关重要的。肠干细胞微环境需要多个信号通路的精确活性梯度来维持肠道的适当稳态。由Clevers团队的ToshiSato博士发表的细胞培养基通过加入细胞因子EGF和R-Spondin1进行环境模拟,以分别激活EGF和WNT通路,并加入Noggin以抑制BMP信号5。
使用肠类器官作为培养系统模型
类器官是一种可用于研究上皮干细胞生物学以及肠结构和功能机制的珍贵模型。虽然这种技术仍处于起步阶段,但已经对各种概念验证进行了相关研究,从而验证其具有广泛的应用。通过DNA转染或siRNA(小干扰RNA),以及使用腺病毒或慢病毒进行病毒感染,可以改变源于小鼠肠隐窝的类器官的基因表达11。 在培养中类器官的快速扩增使其可用于各种分析流程,包括:基因芯片、测序、免疫组织化学和质谱分析12,13。 类器官也可以被冷冻,以便长期储存。最近一些应用到类器官培养物的研究可以分类如下:
干细胞生物学
在开发类器官培养方法不久前,细胞标记和克隆细胞分析让研究人员能够更明确地了解肠隐窝内的干细胞动力学。目前已知隐窝基底部的LGR5+细胞群进行随机对称分裂以维持干细胞池和短暂扩充细胞群6,14。 后续使用类器官的相关研究则确定了嵌入Paneth细胞在干细胞维持中所发挥的复杂作用。去除Paneth细胞即对应于隐窝基底部损失LGR5+细胞15。 Paneth细胞可分泌维持干细胞微环境所必需的细胞因子(如:WNT)。这些细胞被证明对细菌配体、细胞因子和毒蕈碱产生的刺激的敏感性有差异15,16。 当对各细胞类型的具体作用有了更清楚的了解,就可以锁定这些细胞,以治疗因这些细胞异常而造成的疾病,针对炎症性肠病中Paneth细胞突变的治疗就是一个典型的例子。
人类疾病
类器官培养系统集合了多种可成为用于研究人类疾病有用工具的关键属性:首先,患者活检中分离的肠干细胞已生成了类器官17,18;其次,从位于隐窝基底部单个挑选出的EPHB22+细胞可以生成类器官培养物18。 通过基因表达或操作的研究,这类类器官可以用于研究肠疾病的细胞机制。
在许多肠疾病中,对控制增殖和细胞死亡的机制进行抑制具有重要的作用。一些疾病(如:克罗恩氏病、溃疡性结肠炎、腹腔疾病和败血症都会导致免疫系统不受控制地被激活,最终导致肠损伤。应用肠类器官进行的显微镜检查和比色检测法可用于了解上皮细胞存活和细胞死亡的基因功能19。
类器官还为离体了解肠癌的机制提供了一种很有价值的研究工具。缺乏结肠腺瘤性息肉病基因(Adenomatous Polyposis Coli Gene,Apc)的小鼠肠类器官表现出持续性地激活WNT信号,并在注入裸鼠时致瘤,产生高度增殖的管状上皮腺体和明显的基质组织15。 进一步的研究可以详细解释肿瘤形成进程中MAPK和WNT信号通路之间的相互作用。致癌基因BRAF在无蒂锯齿状腺瘤中是最为普遍的突变,其在转基因小鼠中的过度表达将导致MAPK活性的上调。源于携带BRAFV600k突变小鼠的类器官显示总体锯齿状发育不良结构的快速增长,并且显示隐窝区基底部的干细胞池的损失。干细胞损失的程度与WNT/Beta-Catenin的活性呈负相关,这说明可能存在一种“故障-安全”机制,即:MAPK/BRAF信号可以保护肠组织,使其免受致癌活化20。
类器官模型的构架也适用于特定的免疫系统研究。Wilson等通过将细菌病原体直接注射入类器官管腔内而建立起来的肠道感染模型,对Paneth细胞的抗菌反应进行了说明21。 这一技术的发展为研究宿主-微生物的相互作用提供了一种新的模型。
基因治疗
通过特定载体,类器官可以受到病毒的感染。在肠经过人工损伤后,类器官还可以移植到受体小鼠中22。 这两个属性使其可以说明有关修正特定致病突变的问题。由于类器官可以由单个干细胞生成,因此可在转染之后生成克隆类器官并对其进行分析。通过适当组合需要添加的遗传物质可以精确移植类器官,并可以作为基于细胞的一种强有力的治疗工具。最近,有两项将类器官培养物作为完整研究材料之一,侧重于检测开发和后续囊性纤维化跨膜电导受体(CFTR)的基因替代疗法的相关研究。Dekkers等开发了一种基于类器官的检测,通过添加毛喉素,小鼠和人的肠样本均显示由cAMP诱导的野生型类器官的快速膨胀。他们证明,药物引起的快速膨胀情况对于携带CFTR的F508del突变的小鼠会大幅减少,从而建立了一种用于评估针对CFTR突变的基因治疗的新方法23。 Schwank等使用该检测方法,并通过诱导同源重组使用CRISPR/CAS9系统修正了CFTR F508del等位基因。携带修正等位基因的类器官在受到毛喉素刺激时恢复快速膨胀的能力,从而验证了基因替代疗法的原理24。
其他应用
除了考察特定信号通路的研究以外,类器官还为大规模药物筛选提供了研究机会。基于类器官的筛选既可以作为使用CACO-2细胞进行二维筛选的补充,也可以作为一种更具生理相关性的模型系统取代这些基于细胞系的检测25。 目前已进行了利用类器官考察小分子对干细胞生理影响的原理验证性实验。例如,使用可以抑制GSK3Beta和组蛋白脱乙酰基酶的丙戊酸和小分子组合来处理类器官,将导致LGR5+干细胞的大幅增长,并使集落形成效率提高100倍,显示了这些分子在干细胞维持中的作用26。 小分子对野生型和患病类器官产生作用的更多筛选可以成为一种预测性工具,用于扩充目前的ADMET(吸附、分布、代谢、排泄、毒理学)方法论。Spence等证明了类器官的产生不仅限于从原代组织中分离的干细胞。该小组证明了可以通过按时序控制生长因子的操作模拟胚胎内肠发育过程中出现的信号的方法,从而在体外诱导人多能干细胞定向分化为肠类器官。这些人肠类器官不同于直接由肠组织培养的细胞类器官,因为它们含有随上皮一同发育的间充质层,且其发育方式类似于在胚胎发育过程中观察到的方式27。
能够生成与个别患者基因完全相同的类器官,这种能力说明在诊断复杂个别患者疾病方面取得了巨大的进步。类器官培养物可以来自正常和肿瘤人肠道,这一特点使其可以使用这些培养物来筛选特异性肿瘤相关基因突变并制定更加个性化的治疗方案。HUB类器官技术基金(http://hub4organoids.eu/)试图建立一个人体类器官库,或“生物库”,由经过多种筛选方法的匹配健康和患者活检组成。
自第一篇可离体培养肠干细胞的文献发表后,又有多篇文献相继报道了这一技术在未来众多领域中的应用,其中一些已经在本文中进行了介绍。这种新型培养方法取得早期成功的一个关键因素是其能够在多大程度上概括出哺乳动物肠上皮细胞的结构和功能。目前关于类器官技术最令人兴奋的是也可将这种类器官培养方法应用于其它的组织类型。迄今为止,已经建立来自肝脏28、胰腺29、巴雷特食管17、小肠17,18、结肠17,18、大脑3031、前列腺32,33组织,以及来自人诱导多能干细胞27,34,35,36的类器官。在探索新型的移植相关领域研究时,复制这些组织已知功能的能力将使类器官成为一项非常令人兴奋的研究技术。
参考文献
- van der Flier LG and Clevers H. Annu Rev Physiol 71: 241-260, 2009
- Barker N, et al. Nature 449: 1003-1007, 2007
- Buczacki SJA, et al. Nature 495: 65-69, 2013
- Barker N, et al. Genes Dev 22: 1856-1864, 2008
- Sato T, et al. Nature 459: 262-265, 2009
- Snippert HJ, et al. Cell 134-144, 2010
- Sambuy Y, et al. Cell Biol Toxicol 21: 1-26, 2005
- Evans GS, et al. J Cell Sci 101: 219-231, 1992
- Fukamachi H. J Cell Sci 103: 511-519, 1992
- Ootani A, et al. Nat Med 15: 701-706, 2009
- Koo B-K, et al. Nat Methods 9: 81-83, 2012
- Sato T and Clevers H. Science 340: 1190-1194, 2013
- Leushacke M and Barker N. Gut 63: 1345-1354, 2014
- Baker A-M, et al. Cell Rep 8: 940-947, 2014
- Sato T, et al. Nature 469: 415-418, 2011
- Farin HF, et al. J Exp Med 211: 1393-1405, 2014
- Sato T, et al. Gastroenterology 141: 1762-1772, 2011
- Jung P, et al. Nat Med 17: 1225-1227, 2011
- Grabinger T, et al. Cell Death Dis 5: e1228, 2014
- Riemer P, et al. Oncogene, 2014
- Wilson SS, et al. Mucosal Immunol, 2014
- Yui S, et al. Nat Med 18: 618-623, 2012
- Dekkers JF, et al. Nat Med 19: 939-945, 2013
- Schwank G, et al. Cell Stem Cell 13: 653-658, 2013
- Ranga A, et al. Adv Drug Deliv Rev 69-70: 19-28, 2014
- Yin X, et al. Nat Meth 11: 106-112, 2014
- Spence JR, et al. Nature 470: 105-109, 2011
- Huch M, et al. Nature 494: 247-250, 2013
- Huch M, et al. EMBO J 32: 2708-2721, 2013
- Lancaster MA, et al. Nature 501: 373-379, 2013
- Chapman HA, et al. J Clinic Invest 121: 2855-2862, 2011
- Gao D, et al. Cell 159: 176-187, 2014
- Karthaus WR, et al. Cell 159: 163-17520, 2014
- McCracken KW, et al. Nat Protoc. 6(12): 1920-28, 2011
- Watson CL, et al. Nat Med. 20(11): 1310-14, 2014
- McCracken KW, et al. Nature [Epub ahead of print], 2014

 沪公网安备31010102008431号
沪公网安备31010102008431号